Статья:




ПОВЕДЕНЧЕСКИЙ ОТВЕТ САМЦОВ И САМОК БЕЛЫХ КРЫС С РАЗНЫМ УРОВНЕМ АКТИВНОСТИ НА БЛОКИРОВАНИЕ РЕЦЕПТОРОВ ПОЛОВЫХ ГОРМОНОВ
Секция: 2. Биологические науки
Выходные данные
Арчибасова А.В., Чугунова А.Э., Скоропадская А.Н. [и др.] ПОВЕДЕНЧЕСКИЙ ОТВЕТ САМЦОВ И САМОК БЕЛЫХ КРЫС С РАЗНЫМ УРОВНЕМ АКТИВНОСТИ НА БЛОКИРОВАНИЕ РЕЦЕПТОРОВ ПОЛОВЫХ ГОРМОНОВ // Молодежный научный форум: Естественные и медицинские науки: электр. сб. ст. по мат. VI междунар. студ. науч.-практ. конф. № 6(6). URL: https://nauchforum.ru/archive/MNF_nature/6(6).pdf (дата обращения: 28.07.2026)
Лауреаты определены. Конференция завершена
Эта статья набрала 542 голоса
Мне нравится542
Дипломы
лауреатов
лауреатов
Сертификаты
участников
участников
Дипломы
лауреатов
лауреатов
Сертификаты
участников
участников

VI Студенческая международная заочная научно-практическая конференция «Молодежный научный форум: естественные и медицинские науки»
ПОВЕДЕНЧЕСКИЙ ОТВЕТ САМЦОВ И САМОК БЕЛЫХ КРЫС С РАЗНЫМ УРОВНЕМ АКТИВНОСТИ НА БЛОКИРОВАНИЕ РЕЦЕПТОРОВ ПОЛОВЫХ ГОРМОНОВ
Арчибасова Анна Викторовна
студент Донецкого национального университета, Украина, г. Донецк
Чугунова Анастасия Эдуардовна
студент Донецкого национального университета, Украина, г. Донецк
Скоропадская Анна Николаевна
студент Донецкого национального университета, Украина, г. Донецк
Цуканова Евгения Геннадиевна, Хаматшина Екатерина Витальевна
студент Донецкого национального университета, Украина, г. Донецк
Фролова Галина Александровна
научный руководитель, научный руководитель, старший преподаватель Донецкого национального университета, Украина, г. Донецк
Общеизвестным является тот факт, что половые гормоны обладают нейромодуляторным эффектом в отношении некоторых медиаторов в ЦНС [2, с. 45—53; 4, с. 71—78; 6, с. 12—21; 7, с. 29—34]. А поскольку благодаря балансу ряда нейромедиаторных систем мозга, индивид обладает определенным психоэмоциональным профилем, то очевидно, что нарушение обмена половых гормонов может привести к различному комплексу расстройств (например, тревожных и депрессивных) в этой области [3, с. 28—34; 5, с. 4—11]. В связи с широким применением различных препаратов, модулирующих тем или иным образом гормональный статус мужчин и женщин, представляется актуальным исследование влияния данных гормональных систем на психоэмоциональное состояние индивида.
Целью исследования является установление характера влияния блокирования рецепторов половых гормонов у самцов и самок белых крыс с исходно различным уровнем активности на проявления некоторых психоэмоциональных показателей.
Материалы и методы. Эксперимент был проведен на 80 беспородных лабораторных крысах (40 самок и 40 самцов). Поведенческую активность устанавливали в условиях теста продырявленное поле (ПП), маркерным показателем в котором служит исследовательская активность (ИА, суммарное количество вертикальных стоек и заглядываний в отверстия-норки); кроме того, в условиях ПП устанавливали уровень двигательной активности (ДА) как общее количество пересеченных квадратов [7, с. 29—34]. Тревожность определяли в условиях тесте приподнятый крестообразный лабиринт (ПКЛ) по общему времени пребывания на открытом пространстве лабиринта [3, с. 28—34]. Депрессивность и эмоциональность крыс оценивали в тесте Порсолта путем подсчета общего времени неподвижности и количества фекальных болюсов, соответственно [8, с. 53—59]. После контрольного (исходного) тестирования самки и самцы по сигмальному отклонению были разделены на три подгруппы, согласно выраженности у них исследовательского поведения в продырявленном поле. На следующей стадии эксперимента проводилось блокирование рецепторов половых гормонов у животных, после чего крысы проходили повторное тестирование в условиях указанных выше тестов. Блокирование рецепторов андрогенов у самцов проводили путем подкожных инъекций андрофарма (ОАО «Фармак», Украина) в дозе 150 мг/кг в течение 14 дней [5, с. 4—11]. Блокирование рецепторов эстрогенов у самок проводили путем подкожных инъекций тамоксифена (ООО «Фармацевтическая компания «Здоровье», Украина) в дозе 10 мг/кг в течение 14 дней [3, с. 28—34].
Первичные экспериментальные данные обрабатывались с помощью общепринятых методов математической статистики. Разделение исследуемых популяции животных на подгруппы с различным уровнем депрессивности проводилось по сигмальному отклонению. Для оценки достоверности различий между результатами контрольных исследований и для оценки достоверности отличий между опытными и контрольными данными использовался U-критерий Манна-Уитни. Математическая обработка материала проводилась с помощью пакета программ STATISTIKA 6.0 и Excel.
Результаты и их обсуждение. Согласно результатам контрольного (исходного) тестирования в продырявленном поле исходные группы животных разделились на подгруппы с разным уровнем исследовательской активности следующим образом: низкий уровень активности (УА) был показан 35 % самок и 30 % самцов, средний — 40 % и 35 % самок и самцов соответственно, высокий — 25 % от общего числа самок и 35 % от общего числа самок самцов.
Психоэмоциональный профиль самцов и самок крыс, различающихся по уровню исследовательской активности в продырявленном поле в исходных условиях представлен в таблице 1.
Таблица 1.
Психоэмоциональный профиль самок (n=40) и самцов (n=40) с разным уровнем активности в продырявленном поле (контроль), (![]() )
)
|
Поведенческие показатели |
Пол |
Уровень активности |
||
низкий |
высокий |
средний |
||
Исследовательская активность |
♀ |
4,4±0,79## |
15,4±1,15 |
24,2±2,01#●● |
♂ |
4,7±1,09# |
12,6±0,92 |
19,0±0,85#●● |
|
Двигательная активность |
♀ |
9,4±2,36## |
25,3±3,11 |
27,8±1,40●● |
♂ |
8,2±1,09# |
14,0±1,19 |
25,1±2,18#●● |
|
Общее время пребывания на открытом пространстве лабиринта, с |
♀ |
134,6±20,64# |
208,6±15,34 |
222,6±19,35● |
♂ |
76,3±25,13 |
51,6±2,65 |
22,5±9,28# |
|
Суммарное время неподвижности в тесте Порсолта, с |
♀ |
21,7±3,20 |
27,1±4,25 |
26,2±5,34 |
♂ |
17,0±2,77 |
27,3±6,59 |
24,7±4,73 |
|
Уровень эмоциональности |
♀ |
3,8±0,73 |
4,0±1,0 |
0,8±0,22##● |
♂ |
5,3±0,49 |
4,7±0,81 |
5,4±0,48 |
|
Примечание: #, ## — различия статистически значимы (р<0,05) и (р<0,01) соответственно в сравнении показателей условного контроля (средний уровень активности) с группами высокого и низкого уровней активности; ●, ●● — различия статистически значимы (р<0,05) и (р<0,01) соответственно при сравнении показателей группы с крайними типами выраженности активности.
Как видно из данных, представленных в таблице, у средне- и высокоактивных в исходных условиях самок уровень выраженности двигательной активности не отличается. У самцов же данный показатель четко коррелирует с уровнем исследовательской активности. Обращает на себя внимание тот факт, что уровень депрессивности обоих полов, устанавливаемый в тесте Порсолта, одинаков у всех животных не зависимо от исходного уровня их активности. Аналогичным образом, не различаются по уровню эмоциональности самцы разных подгрупп.
Однако, выявлено, что исходно низкоактивным самкам и высокоактивным самцам свойственна большая тревожность, нежели другим подгруппам крыс, а исходно высокоактивным самкам — меньшая эмоциональность.
На следующем этапе исследования устанавливалась степень влияния блокирования рецепторов андрогенов у самцов и эстрогенов у самок на выраженность некоторых психоэмоциональных характеристик у крыс с исходно разным уровнем активности. Так, установлено, что блокирование рецепторов половых гормонов привело к угнетению проявлений как исследовательской, так и двигательной активности у высокотревожных самок и самцов в среднем на 60—67 % (p<0,01) в случае с ИА и 50—58 % (p<0,01) в случае с ДА (рис. 1, 2). Антиэстрогенное воздействие тамоксифена у самок с исходно средним уровнем активности привело к аналогичному эффекту — значительному сокращению проявлений поведенческой активности (на 85—90 %, p<0,01). У самцов же этой подгруппы блокирование рецепторов андрогенов привело к увеличению двигательной активности. Исходно низкоактивные крысы чувствительности к данным фармакологическим воздействиям в условиях продырявленного поля не проявили.
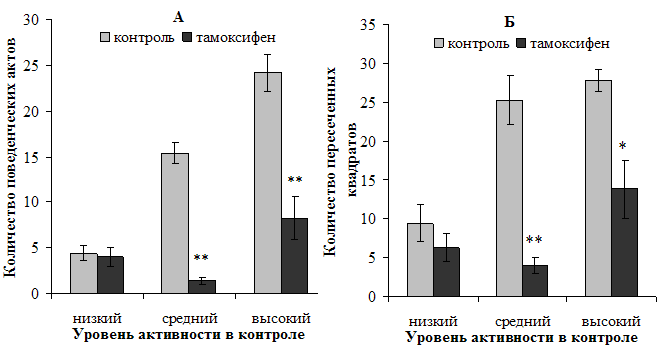
Рисунок 1. Характер влияния тамоксифена на исследовательскую (А) и двигательную (Б) активность самок в продырявленном поле
Примечание: *, ** — различия статистически значимы (р<0,05) и (р<0,01) соответственно при сравнении с исходными показателями.
Относительно влияния блокирования рецепторов половых гормонов на показатель тревожности крыс в приподнятом крестообразном лабиринте, отличающихся по исходному уровню активности, установлено следующее. Антиэстрогенное действие тамоксифена оказало анксиогенный эффект на высокоактивных самок: время пребывания на открытом пространстве у крыс данной подгруппы сократилось на 63,6 % (p<0,01) относительно исходных значнений (рис. 3, А). На тревожность самок остальных подгрупп тамоксифен не повлиял. У самцов же блокирование рецепторов половых гормонов оказало анксиолитический эффект, проявившийся в увеличении маркерного показателя ПКЛ на 143,8 % (p<0,01) и 700 % (p<0,01) у средне- и высокоактивных крыс соответственно (рис. 3, Б).

Рисунок 2. Характер влияния андрофарма на исследовательскую (А) и двигательную (Б) активность самцов в продырявленном поле
Примечание: *, ** — различия статистически значимы (р<0,05) и (р<0,01) соответственно при сравнении с исходными показателями.
Ярко выраженный депрессогенный эффект блокирования рецепторов эстрогенов был выявлен у самок всех исходных подгрупп активности в условиях теста Порсолта (рис. 4, А): увеличение показателя депрессивности составило в среднем от 55 до 70 % (p<0,05). На депрессивный статус самцов андрофарм оказал антидепрессивный эффект, однако достоверными эти изменения не оказались (рис. 4, Б).
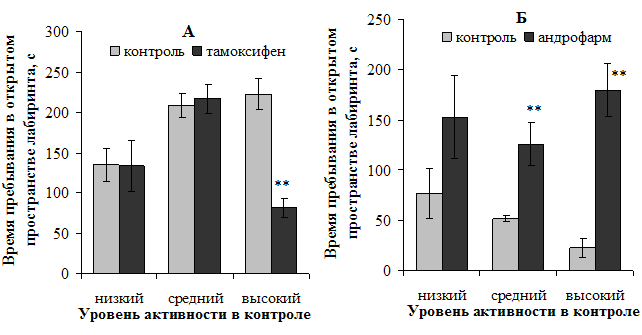
Рисунок 3. Характер влияния блокирования рецепторов половых гормонов на время пребывания на открытом пространстве самок (А) и самцов (Б) в приподнятом крестообразном лабиринте
Примечание: ** — различия статистически значимы (р<0,01) при сравнении с исходными показателями.
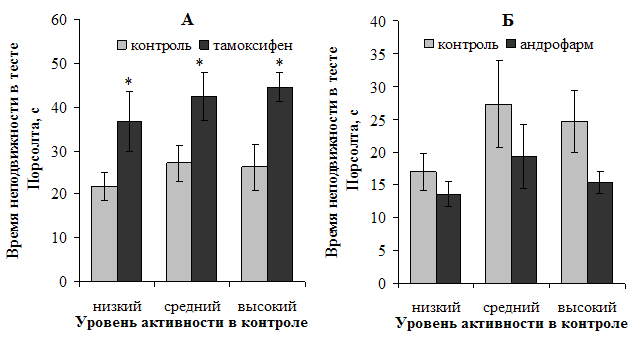
Рисунок 4. Характер влияния блокирования рецепторов половых гормонов на показатель депрессивности самок (А) и самцов (Б) в тесте Порсолта
Примечание: * — различия статистически значимы (р<0,05) при сравнении с исходными показателями.
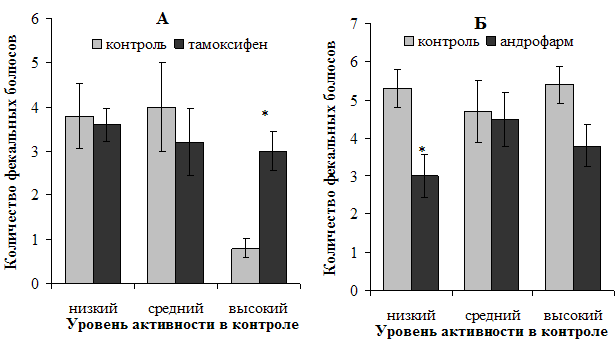
Рисунок 5. Характер влияния блокирования рецепторов половых гормонов на показатель эмоциональности самок (А) и самцов (Б) в тесте Порсолта
Примечание: * — различия статистически значимы (р<0,05) при сравнении с исходными показателями.
Характер влияния используемых препаратов на эмоциональность животных представлен на рисунке 5. Очевидно, что тамоксифен оказал влияние на эмоциональность только высокоактивных в контроле самок в сторону ее увеличения (в 3,8 раз, p<0,05); действие андрофарма же привело к угнетению проявлений данного показателя на треть (p<0,05) у исходно низкоактивных самцов.
Выводы:
1. Результаты исходного тестирования исследуемых животных в тесте продырявленное поле позволили выделить среди однородной популяции крыс животных, отличающихся по уровню активности. Учитывая, что экспериментальные животные с момента рождения содержались в одинаковых условиях и получали одинаковую пищу, проведенные эксперименты позволили сделать вывод, что такой психоэмоциональный показатель как уровень активности является генетически детерминированным.
2. Животные, отличающиеся по исходному уровню активности не отличаются по уровню депрессивности.
3. Самки и самцы с исходно низким уровнем активности не проявляют чувствительности к блокированию эстрогеновых и андрогеновых рецепторов соответственно.
4. Избирательное блокирование рецепторов эстрогенов у самок оказывает депрессогенный эффект на животных не зависимо от исходного уровня их поведенческой активности. Избирательное блокирование рецепторов андрогенов у самцов оказывает анксиолитический эффект на животных не зависимо от исходного уровня их поведенческой активности.
Список литературы:
- Амстиславская Т.Г. Половая активация самцов: поведение и гормональный ответ / Т.Г. Амстиславская, К.В. Осипов // Бюллетель СО РАН. — 2003. — № 3. — с. 112—114.
- Бабичев В.Н. Влияние эстрогенов на центральную нервную систему / В.Н. Бабичев // Вестн. Рос.АМН. — 2005. — № 6. — с. 45—53.
- Казакова С.Б. Влияние тамоксифена на тревожность у интактных и овариоэктомированных самок крыс / С.Б. Казакова, Ю.О. Федотова, Н.С. Сапронов // Экспер. и клинич. фармакология. — 2007. — № 5. — с. 28—34.
- Караева Е.Н. Новые аспекты действия эстрогенов / Е.Н. Караева // Экспер. и клинич. фармакология — 2003. — № 4. — с. 71—78.
- Резников А.Г. Блокаторы рецепторов андрогенов и их применение в биологии и медицине / А.Г. Резников // Досягнення біології та медицини — 2004. — № 1. — с. 4—11.
- Сапронов Н.С. Взаимодействие нервных и гормональных факторов в реализации высших функций мозга / Н.С. Сапронов, Ю.О. Федотова, О.О. Масалова // Мед.академич. журн. — 2008. — Т. 8, № 1. — с. 12—21.
- Сапронов Н.С. Половые гормоны и поведенческие реакции. / Н.С. Сапронов, Ю.О. Федотова, Н.П. Гончаров // Вестник Рос. АМН. — 2001. — № 12. — с. 29—34.
- Porsolt R.D. Animal models of depression. Utility for transgenic research // Rev. Neurosci. 2000. — № 11. — Р. 53—59.
