Статья:




Особенности цито- и хемоархитектоники энторинальной коры мозга плода человека во втором триместре гестации
Секция: Биология
Выходные данные
Моисеенко И.А., Андреева Д.Д., Аксенова Л.Е. [и др.] Особенности цито- и хемоархитектоники энторинальной коры мозга плода человека во втором триместре гестации // Молодежный научный форум: Естественные и медицинские науки: электр. сб. ст. по мат. XLII междунар. студ. науч.-практ. конф. № 2(41). URL: https://nauchforum.ru/archive/MNF_nature/2(41).pdf (дата обращения: 29.07.2026)
Лауреаты определены. Конференция завершена
Эта статья набрала 0 голосов
Мне нравится0
Дипломы
лауреатов
лауреатов
Сертификаты
участников
участников
Дипломы
лауреатов
лауреатов
Сертификаты
участников
участников

XLII Студенческая международная заочная научно-практическая конференция «Молодежный научный форум: естественные и медицинские науки»
Особенности цито- и хемоархитектоники энторинальной коры мозга плода человека во втором триместре гестации
Моисеенко Ирина Александровна
аспирант каф.эмбриологии биологического факультета Санкт-Петербургского Государственного университета (СПбГУ), РФ, г. Санкт-Петербург
Андреева Дарья Дмитриевна
студент 3 курса бакалавриата каф.цитологии и гистологии биологического факультета Санкт-Петербургского Государственного университета (СПбГУ), РФ, г. Санкт-Петербург
Аксенова Любовь Евгеньевна
студент 3 курса каф.цитологии и гистологии бакалавриата биологического факультета Санкт-Петербургского Государственного университета, (СПбГУ), РФ, г. Санкт-Петербург
Сидорова Надежда Александровна
аспирант каф. патологической анатомии с курсом судебной медицины Санкт-Петербургского государственного педиатрического медицинского университета (СПбГПМУ), РФ, г. Санкт-Петербург
Зыкин Павел Александрович
научный руководитель, канд. биол. наук, доц. кафедры цитологии и гистологии СПбГУ, РФ, Санкт-Петербург
Работа выполнена по гранту СПбГУ №1.38.333.2015, на оборудовании РЦ «РМиКТ» Научного Парка СПбГУ.
Энторинальная кора (ЭК) – относится к межуточной, отделяющей архи- и палеокортикальные формации от неокортекса, и входит в состав медиальной темпоральной доли (MTL). На сегодняшний день ЭК рассматривают как интерфейс между сенсорными, ассоциативными областями неокортекса и гиппокампальными формациями [2]. Несмотря на свои небольшие размеры, ЭК является структурно и функционально неоднородным образованием, её наиболее крупные субструктуры – медиальная (МЭК) и латеральная (ЛЭК) области происходят из разных прогениторных зон. По экспрессии 7 генов (Lef1, Lhx2, Lhx9, Lhx5, Lmo3, Lmo4 и Prox1) в мозге мышей доказано, что МЭК и гиппокамп происходят из медиального паллиума, а ЛЭК – из дорзо-латерального [1]. Функциональные различия МЭК и ЛЭК у животных заключаются в том, что нейроны МЭК и дорзального гиппокампа обрабатывают информацию о пространственных стимулах, а нейроны ЛЭК и вентрального гиппокампа обеспечивают распознавание объектов и реакцию на новизну [7]. Благодаря совершенствованию метода функциональной магнито-резонансной томографии (фМРТ) к настоящему времени установлено, что у человека, также как и у животных, в формировании пространственной памяти задействована МЭК, а предметной – ЛЭК [4]. Сведения о цитоархитектонических полях ЭК мозга человека весьма разноречивы: методами цитоархитектники выделяют от 7 до 28 полей, в соответствии с нейрохимической характеристикой нейронов выделено восемь цитоархитектонических полей ЭК [6]. Тщательное исследование характеристик ЭК значимо не только для понимания нейронных механизмов человеческой памяти, но имеет важное клиническое значение. Наиболее очевидные слой- и регионально- специфичные изменения известны для болезни Альцгеймера, при которой наиболее выраженно страдает слой II МЭК, при смешанной деменции – страдает слой III, при болезни Гентингтона –слой VI [6]. Очень многое в организации ЭК мозга человека, её полей и слоёв может раскрыть плодный период, когда гирификация коры только начинается, а ЭК более доступна для исследования. Этот факт констатируется практически всеми исследователями этой корковой области, но такие работы единичны и ограничиваются, в основном, изучением цитоархитектоники. Наиболее полной можно считать работу G.Macci [5], выполненную методами цито- и миелоархитектоники. В настоящий момент наиболее важные нерешенные вопросы касаются хемоархитектоники, нейронной организации слоёв МЭК и ЛЭК, закономерностей их онтогенеза.
Цель настоящей работы заключается в изучении цито- и хемоархитектоники энторинальной коры мозга плодов человека второго триместра гестации.
Материал и методы исследования
В работе использовали 5 левых полушарий мозга плодов в возрасте 20–24 недель гестации, полученных в результате легальной аутопсии в Санкт-Петербургском педиатрическом медицинском университете, и с разрешением на последующее исследование этического комитета СПбГПМУ. Весь материал, с постмортемным сроком в пределах 24 часов, по заключению патологоанатома не имел патологий и кровоизлияний, показал отрицательную реакцию на вирусы нейроинфекций: герпеса (тип 1), цитомегаловируса, Эпштейна-Барр. Материал фиксировали в 4% растворе параформальдегида на 0,1М фосфатном буфере, РН 7,4. Выделенные блоки височной области коры заливали в парафин, фронтальные срезы толщиной 12 мкм изготавливали на микротоме Leica RM 2245. Для последующего исследования срезы окрашивали крезиловым фиолетовым по Нисслю или подвергали иммуногистохимической (ИГХ) обработке с антителами к белкам цитоскелета (МАР2 и N200), рилину, кальций-связывающим белкам (парвальбумину и калретинину). Исследование препаратов проводили с помощью микроскопов Leica DM 5500 и Leica TCS SP5, оборудованных автоматизированными предметными столиками и фотокамерами. Часть материала обрабатывали по методу CLARITY, который позволяет исследовать очень толстые, до 500 мкм, срезы мозга. Выделенные блоки коры пропитывали акриламидным гидрогелем, который связывает белки, но не липиды [3]. Последние удалялись из ткани, вследствие чего блок становился гораздо более прозрачным. Впоследствии такие блоки подвергали ИГХ обработке с антителами к вышеуказанным белкам и микроскопировали на конфокальном микроскопе Leica TCS SP5.
Результаты исследования.
У плодов 20–24 недель гестации энторинальная кора прослеживается от рострального полюса височной доли, а каудально до уровня латерального коленчатого тела. Уже на 20 неделе развития ЭК по характеру и степени стратификации отличается от пограничных областей новой коры, гиппокампальных структур и периамигдалярной области. Слой I характеризуется редкоклеточность, и содержит крупные нейроны. При ИГХ обработке эти нейроны демонстрируют рилин-позитивность, что позволяет идентифицировать их как клетки Кахаля-Ретциуса. Слой II в МЭК образуют регулярно расположенные, характерные именно для этой области, островки плотно упакованных МАР2-позитивных клеток. Слой III заметно расширяется от МЭК к ЛЭК, содержит МАР2-позитивные клетки только в ЛЭК. Слой IV – свободная от нейронов диссиканта, является характерной особенностью ЭК. Под ней располагается единый комплекс клеток, который трудно разделить на слои V и VI (рис. 1), в нём присутствует небольшое количество МАР2-позитивных пирамидных нейронов. Исследование блоков ткани, обработанных по методу CLARITY и ИГХ с антителами к N200 и калретинину, позволило более детально изучить организацию слоя I. В его составе были обнаружены крупные N200-позитивные клетки Кахаля-Ретциуса с характерным для этого типа нейронов ветвлением дендритов и N200-позитивное сплетение тангенциальных волокон. По нашему предположению это сплетение входит в состав перфорантного тракта. Калретинин-позитивные элементы были приурочены к телам и дендритам клеток Кахаля-Ретциуса, что позволило нам обоснованно предположить, что они являются пресинапсами окончаний, приуроченных к этим нейронам (рис. 2). Полученные результаты дополняют ранее полученные сведения об организации маргинальной зоны коры мозга человека в плодный период [8].
Вывод: комплексное использование цитоархитектонических и ИГХ методов, дополненных CLARITY, позволяет разделить энторинальную кору мозга плодов человека второго триместра гестации на медиальную и латеральную области, а также получать сведения о нейронной организации этой корковой формации.
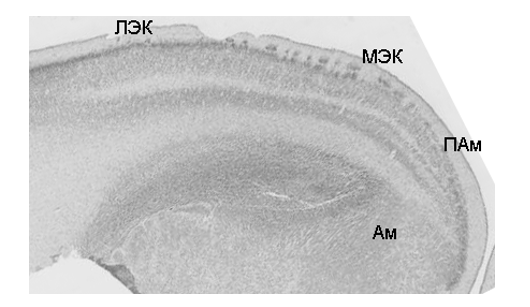
Рисунок 1. Цитоархитектоника энторинальной коры мозга плода человека 22 недель гестации. Обозначения: ЛЭК, МЭК – латеральная и медиальная энторинальная кора, ПАм – периамигдалярная область, Ам – амигдала. Окраска по Нисслю. Ув.х5,5

Рисунок 2. Проекция слоя I энторинальной коры мозга плода человека 24 недель гестации, препарат изготовлен по методу CLARITY. Обозначения: К-Р – клетки Кахаля-Ретциуса, ТВ – сплетение тангенциальных волокон
Список литературы:
1. Abellán A., Desfilis E., Medina L. Combinatorial expression of Lef1, Lhx2, Lhx5, Lhx9, Lmo3, Lmo4, and Prox1 helps to identify comparable subdivisions in the developing hippocampal formation of mouse and chicken//Front. Neuroanat. 2014. doi: 10.3389/fnana.2014.00059.
2. Canto C.B., Wouterlood F.G., Witter M.P. What does the anatomical organization of the entorhinal cortex tell us? // Neural Plasticity. 2008. doi: 10.1155/2008/381243.
3. Chung K., Deisseroth K. CLARITY for mapping the nervous system. Nature Methods.// 2013.10 (6), 508–513.
4. Maass A., Berron D., Libby L., Ranganath, C., Düzel E. Imaging the human medial temporal lobe with high-resolution fMRI//Neuron. 2015. 65, 298–308.
5. Macci G., The ontogenetic development of the olfactory telencephalon in man // J. Comparative Neurology. 1951. 95 (1), 245–305.
6. Mikkonen M., Alafuzoff I., Tapiola T., Soininen H. and Miettinen R. Subfield- and layer-specific changes in parvalbumin, calretinin and calbindin-d28k immunoreactivity in the entorhinal cortex in Аlzheimer’s disease//Neuroscience. 1999. 92 (2), 515–532.
7. Sasaki T., Leutgeb S., and Leutgeb J.K. Spatial and memory circuits in the medial entorhinal cortex//Curr Opin Neurobiol. //2015. doi:10.1016/ j.conb. 2014.10.008.
8. Tkachenko L.A. Zykin P.A, Nasyrov R.A. Krasnoschcekova E.I. Distinctive features of the human marginal zone and Сajal–Retzius cells: comparison of morphological and immunocytochemical features at midgestation. // Front. Neuroanat. 2016. doi: 10.3389/fnana.2016.00026.
