Биоэкологические особенности Rhaponticum integrifolium C. Winkl. в Узбекистане
Конференция: XXII Международная научно-практическая конференция «Научный форум: медицина, биология и химия»
Секция: Ботаника

XXII Международная научно-практическая конференция «Научный форум: медицина, биология и химия»
Биоэкологические особенности Rhaponticum integrifolium C. Winkl. в Узбекистане
Bioecological features of Rhaponticum integrifolium C. Winkl. on the territory of Uzbekistan
Namuna Aliyeva
Senior lecturer, Kokand state pedagogical Institute, Uzbekistan, Kokand
Аннотация: В статье приведены данные о биоэкологических особенностях Rhaponticum integrifolium, содержащего экдистероид, произрастающего в разных экологических условиях на территории Узбекистана. Выявлено что, продолжительность жизненного цикла Rh. Integrifolium, произрастающего на территории кишлаков Гилан и Чодак близка (17-18 лет) без существенных различий в смене периодов и этапов онтогенеза. Однако, для растений, произрастающих на территории кишлака Чодак, характерно усиленное развитие надземных частей, а на территории Гилан активное развитие подземных частей. Продолжительность фенологических фаз варьирует от 100-115 дней (Чодак) до 125-135 дней (Гилан), что свидетельствует о чувствительности развития растения к температуре воздуха, влажности почвы и высоте над уровнем моря. С приближением к средневозрастному состоянию у растений наблюдается относительно раннее начало фенологических фаз и это связано с характерными биологическими особенностями растения.
Abstract. The article presents data on bioecological features of ecdysteroid-containing plants Rhaponticum integrifolium in two conditions of Uzbekistan. As a result, it is revealed that, the duration of a large Rh life cycle. integrifolium in the territories of Gillan and Chodak is close (17-18 years) to each other and there are no special differences in the change of periods and stages. However, for plants growing on the territory of Chodak, characterized by increased development of above-ground parts, and on the territory of Gilan active development of underground parts. 5. The duration of phenological phases varies from 100-115 days (Chodak) to 125-135 days (Gillan), indicating the sensitivity of the seasonal development of the plant to air temperature, soil moisture and altitude. With the approach to the middle age state of the plant there is a relatively early onset of phenological phases and this is due to the characteristic biological characteristics of the plant.
Ключевые слова: Rh. Integrifolium; онтогенез; рост и развитие.
Keywords: Rh. Integrifolium; ontogenesis; growth and development.
В мире при лечении опасных для здоровья человека заболеваний применяется более 3000 видов средств естественного происхождения и 35% из них составляет растительное сырье. Поэтому, широкомасштабное применение экдистероидов в современной медицине требует определение видов растений, содержащих их производные и оценки состояния их сырья. В связи с этим, изучение видов растений, содержащих производные экдистероидов, из локальной флоры, выявление их биоэкологических особенностей и внедрение в производство имеет важное научно-практическое значение.
На сегодняшний день в мире экдистероид-содержащие растения не выращиваются в масштабе удовлетворяющем потребности фармацевтической промышленности. Из многочисленных ботанико-морфологических и экологических исследований в различных климатических условиях выявлено, что для производства экдистероид-содержащих препаратов интродукция видов рода Rhaponticum, содержащих в своем составе в большом количестве фитоэкдистероиды, эффективно с экономической точки зрения [12].
В современной научной медицине экдистероиды природного происхождения применяются при нарушениях сердечно-сосудистой, центральной нервной и репродуктивной систем. Кроме того, используются при умственном и физическом утомлении в качестве бодрящего и стимулирующего средства, при потере работоспособности, при импотенции и ослаблении функций различных органов [7], [9].
В составе 95% представителей флоры всего мира присутствуют следы экдистероидов, а у 4-6% они содержатся в количестве 0,001-0,0001%, и лишь у считанных видов в определенных фазах развития в составе некоторых надземных органов количество экдистероидов может составлять 1-3 % [3], [13], [17], [8].
Род Rhaponticum включает в себя 17 видов, распространенных в узком и почти сплошном ареале лентовидной формы в Северном полушарии от Атлантического океана до берегов Тихого океана. Только 1 вид распространен в Австралии [2]. На территории Средней Азии встречаются 9 видов рода Rhaponticum [2], [4].
Впервые наличие экдистероидов у представителей рода Rhaponticum выявлено и выделено учеными Н.К. Абубакировым, У. Болтаевым и Ш.Г. Ганиевым. На сегодняшний день 9 видов их используются в фармацевтике и медицине [1], [6].
Rh. integrifolium встречается в Западно Памиро-Алайских горах – в районах Зерафшанского, Гиссарского, Картегинского, Дарвазского, Бабатауского и Ферганского хребтов, в районе хребта Петра Первого и Южно-Таджикистанских низкогорях. В надземных органах растения содержится 0,3% экдистерона, в стеблях – 0,1% и в листях – 0,1% [1], [6], [8], [10], [14], [15], [16].
Учеными Института химии растительных веществ АН РУз доказана биэквивалентность экдистерона, выделенного из надземной части Rh. integrifolium с экдистероном сырья Rh. carthamoides, привезенного из России [5], [11].
Из анализа литературных источников можно сказать, что кроме изучения биохимического состава, ресурсов и распространения в государствах СНГ разработаны и усовершенствованы технологии размножения и методы выращивания Rh. carthamoides. в различных почвенно-климатических условиях. Однако, на сегодняшний день биоэкологические особенности Rh. integrifolium в Узбекистане изучены недостаточно. Исходя из этого, изучение онтогенеза и семенной продуктивности растения, обоснование изменчивости количества экдистероидов в различных частях растения, оценка роли растения в растительном покрове и природных ресурсах приобретают важное научно-практическое значение.
В связи с этим, целью исследования явилось обоснование биоэкологических особенностей и оценка природных ресурсов Rhaponticum integrifolium в условиях Узбекистана.
В результате исследований выявлено, что размеры семян распространенных в различных географических широтах и долготах растений Rh. intеgrifolium резко отличаются друг от друга (в районе Чодак: длина 0,9±0,25 см, ширина 0,25±0,11 см, масса 1000 семян 22,5±0,67 гр; в тайоне Гилан: длина в среднем 1,1±0,15 см, ширина 0,37±0,13 см, масса 1000 семян 23,5±0,94 гр).
В лабораторных условиях повышение температуры с 180C до 280C положительно влияет на всхожесть семян Rh. integrifolium. Выявлено, что всхожесть семян проявляет чувствительность к процессу стратификации (27-33%). В полевых условиях максимальная всхожесть семян Rh. integrifolium наблюдается при посадке в октябре-ноябре после механической обработки и хранении в холодильнике (68-75%). В подобных условиях семена прорастают на 20-25 дней раньше по сравнению с необработанными семенами в благоприятных погодных условиях и образуют корневую систему до повышения температуры воздуха и осушения 10-20 см горизонта почвы.
Виргинильный период. 1-год. Этап проростка (p). Прорастание семян Rh. integrifolium надземное, из семени прорастает первичный (главный) корень. После достижения длины корня 1,5-2 см быстроразвивающиеся семядольные листья поднимаются над поверхностью почвы. Длина новых семядольных листьев 1,5-2 см, ширина – 0,6-0,7 см.
У впервые проросших растений к 20-25 апрелю образуются настоящие листья в середине семядольных листьев. К этому времени корневая система проростка формируется и достигает 6-9 см. В естественных условиях выживают 2-3 % проростков. Однако, на опытных участках при помощи соответствующих агротехнических мер данный показатель достигает 80-85 %.
Ювенильный этап (j). Растение на ювенильный этап развития переходит через 20-24 дней. Данный этап характеризуется образованием 6-10 прикорневых листьев, появлением нового подземного органа – стеблекорня, активным формированием корней и образованием спящих почек у корня. В начале этапа спустя 3-5 дней после образования первого настоящего листа появляются вторые настоящие листья. Последующие образующиеся листья Rh. Integrifolium, по сравнению с предыдущими, отличаются крупностью черешка и листовой пластинки и ускоренным развитием.
На ювенильном этапе одним из важных морфологических изменений, наблюдаемых у растения, является утолщение гипокотиля и формирование придаточных корней, образующихся во внутренних тканях. При этом формируется подземный орган – стеблекорень с новым функциональным строением. На ювенильном этапе в надземной части стебля, в нижней центральной части черешков листьев образуются 2‑6 конусовидных почек – разовьющихся весной следующего года.
Вегетация Rh. integrifolium продолжается с конца июля до первой декады августа и заканчивается с образованием у каждой особи до 6‑7 прикорневых листьев.
В ходе исследований в первый год вегетации у Rh. integrifolium не наблюдались признаки, свойственные имматурному этапу. По нашему мнению, отсутствие имматурного этапа в условиях Узбекистана на первом году вегетации связано с проявлением морфо-биологических приспособлений к аридным условиям территории с ускорением этапов онтогенеза растения в жарких погодных условиях. Подобное морфобиологическое приспособление проявляется также в виде активности надземных и подземных частей растений, прорастающих в условиях Чодак (1000-1200 м над ур.м) и Гилан (2200-2400 м над ур.м). Растения произрастающие на опытных участках Гилан отличаются активным развитием подземных органов, а произрастающие на территории Чодак – активным развитием надземных органов – листьев, кроме того количеством восстанавливающихся почек.
2-год. Виргинильный этап (v). В большом жизненном цикле Rh. integrifolium виргинильный или зрелый этап продолжается до 3 лет (Таблица 1). Этап характеризуется полным формированием вегетативных органов, повышением биомассы, формированием настоящих розеточных листьев из стеблекорня и сильным развитием корневой системы (Рисунок 1).

Рисунок 1. Этапы онтогенеза Rhaponticum integrifolium
Появление клубневидных расширений на уровне 12-15 см основного корня на третьем году виргинильного этапа имеет особое значение. Клубневидная часть корня сохраняется до последнего года виргинильного этапа и на 2-этапе генеративного периода исчезает в результате усиленного развития главного корня.
Генеративный период. Молодые генеративные растения (g1). Первый почечный побег Rh. integrifolium переходит в генеративный период на втором году вегетации. Если сравнить по количеству особей, то на третьем году вегетации 7-8 % растений переходят в генеративный период, на четвертом году – 75-80 %, на пятом году вегетации – 10-12 % (Таблица 1).
Большинство генеративных побегов, образованных на третьем году вегетации, не проходят полный цикл развития. У большинства особей наблюдается ослабление развития междоузлий на начальных этапах развития генеративных побегов, что приводит к высыханию соцветий. Лишь незначительное количество генеративных побегов – 10-15 % могут достигать фазы цветения. На опытных участках в районах Гилан и Чодак на 3-4 год вегетации на каждом растении формируется по 1 генеративному побегу. Если длина образованного в первом году вегетации генеративного побега невысока (20-28 см), то в последующих годах превосходит (70-90 см) вегетативного побега (Таблица 1). Одной из основных особенностей первого генеративного этапа является увеличение количества запасающих почек. Их увеличение наблюдается, особенно, на 4 году вегетации растений, что обеспечивает переход растений к зрелому генеративному этапу (g2).
Средневозрастные генеративные растения (g2). Rh. integrifolium с 5 года вегетации переходит на зрелый генеративный этап, который продолжается до 10 года вегетации. На 5-год вегетации 40-45 % растений, а на 6-7 год вегетации 100% растений переходят в полную зрелую генеративную стадию развития. При этом, может образоваться до 6-10 генеративных побегов. В 2-4 верхних узлах центрального генеративного побега наблюдается образование 2 генеративных веточек и формирование в них дополнительных соцветий. Однако, образованные в центральном генеративном побеге соцветия отличаются более меньшим объёмом от остальных соцветий (Таблица 1, рисунок 1). Несмотря на максимальное значение всех морфологических параметров Rh. integrifolium, на зрелом генеративном этапе начинается формирование партикул, определяющий переход растения на сенильный период. На g2 этапе 10-15 % растений образует партикулы.
Таблица 1
Изменение морфометрических показателей Rhaponticum integrifolium в виргинильном (v), генеративном (g1, g2, g3) и сенильном (ss, s) периодах, Гилан/Чодак
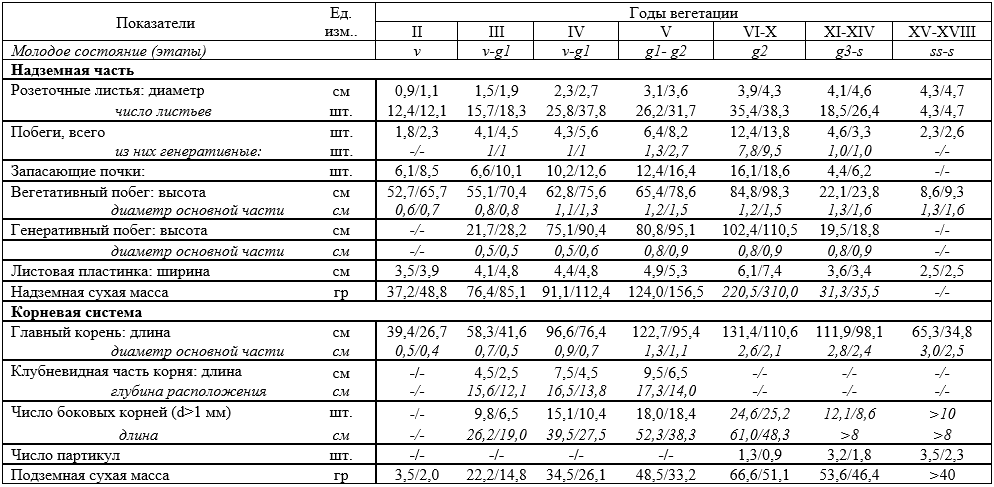
Старые генеративные растения (g3). К 9, и особенно 10-11 году вегетации у Rh. integrifolium ослабляется формирование вегетативных и генеративных органов. Уменьшение количества розеточных листьев, запасающих почек, генеративных побегов, уменьшение объема соцветий и листовой пластинки и самое главное – гниение корня свидетельствуют о переходе растения на старый генеративный этап. На данном этапе Rh. integrifolium начинает свою вегетацию на 15-20 дней позже по сравнению с ранневегетирующими растениями и образование генеративных побегов осуществляется за счет старых генеративных полициклических побегов. Их число не превышает 1 шт. и высота намного ниже (18-20 см), чем вегетативные побеги (Таблица 1, рисунок 1).
В старом генеративном возрасте формирование партикул ускоряется и составляет 2-3 шт. У Rh. integrifolium старый генеративный этап продолжается 3-4 года. Несмотря на ослабление всех морфологических признаков, растения на этом этапе отличаются увеличением диаметра розетки надземной части и диаметра основы главного корня, к тому же эта тенденция продолжается почти до конца сенильного периода.
Сенильный период. У Rh. integrifolium предшествующий сенильному периоду (s) субсенильный этап (ss) начинается на 13-14 году вегетации и ускоряется на 15 году вегетации. При этом, габитус растения существенно уменьшается, розеточная часть утолщается за счет партикул и верхняя часть главного корня также укрупняется. 2/3 длины главного корня исчезает (Таблица 1, рисунок 1). Обычно Rh. integrifolium свою вегетацию заканчивает на 17-18 году вегетации, однако некоторые растения погибают в возрасте 20-21 года.
Подводя итоги следует отметить, что продолжительность большого жизненного цикла Rh. integrifolium на территориях кишлаков Гилан и Чодак близка друг к другу и не отмечено заметных отличий при смене периодов и этапов развития. Однако, выявленные различия морфологических показателей в онтогенезе связано с почвенно-климатическими условиями территорий.
Онтогенез Rh. integrifolium продожается 17-18 лет. Первый год вегетации включает в себя стадии проростка и ювенильный этап виргинильного периода. В 1-2 годы жизни растения отсутствует имматурная стадия и на 2 год вегетации Rh. integrifolium переходит на виргинильный этап. Отсутствие имматурного этапа в условиях Узбекистана связано с проявлением морфо-биологических приспособлений к аридным условиям территории с ускорением этапов онтогенеза растения в жарких погодных условиях. Виргинильный этап растения продолжается 3 года, но с 3 года вегетации начинается генеративный период. Генеративный период растения охватывает 10-11 лет или почти 60% онтогенеза. Старость растения начинается с 13-14 лет и продолжается до 17-18 лет. Для растений, произрастающих на территории Чодак, характерно усиленное развитие надземных частей, а для растений, произрастающих на территории Гилан активное развитие подземных частей.
В исследованиях выявлено, что общая вегетация Rh. integrifolium на опытном участке Гилан равна 125-135 дней, а на опытном участке Чодак – 100-115 дней. Для фазы бутонизации потребуется 10-15 дней вегетации, для цветения 24-26 дней (на территории Чодак 20-21 дней), для формирования семян 30-35 дней (на территории Чодак – 25-30 дней), для созревания семян 25-32 дней (на территории Чодак 20-22 дней).
Начало, конец и продолжительность фенологических фаз растения связаны с температурой воздуха, влажностью почвы и с периодами онтогенеза. Повышение температуры воздуха на относительно равнинных территориях, расположенных на высоте 1000-1200 м над ур. моря влияет на сокращение фенологических фаз (территория Чодак) или влажная погода и относительно низкая температура на высоте 2200-2400 м над ур. моря влияет на продолжительность фенологических фаз (территория Гилан). Кроме того, с приближением к средневозрастному периоду у растений наблюдается относительно раннее начало фенологических фаз и это, вероятно, связано с специфическими биологическими особенностями растения.
