Статья:




Активность NА,К-АТФазы различных отделов головного мозга крыс, подвергнутых действию иммобилизационного стресса и обработанных антихолинэстеразным препаратом
Секция: Биология
Выходные данные
Абрамова Н.С. Активность NА,К-АТФазы различных отделов головного мозга крыс, подвергнутых действию иммобилизационного стресса и обработанных антихолинэстеразным препаратом // Молодежный научный форум: Естественные и медицинские науки: электр. сб. ст. по мат. XLV междунар. студ. науч.-практ. конф. № 5(45). URL: https://nauchforum.ru/archive/MNF_nature/5(44).pdf (дата обращения: 27.07.2026)
Лауреаты определены. Конференция завершена
Эта статья набрала 0 голосов
Мне нравится0
Дипломы
лауреатов
лауреатов
Сертификаты
участников
участников
Дипломы
лауреатов
лауреатов
Сертификаты
участников
участников

XLV Студенческая международная заочная научно-практическая конференция «Молодежный научный форум: естественные и медицинские науки»
Активность NА,К-АТФазы различных отделов головного мозга крыс, подвергнутых действию иммобилизационного стресса и обработанных антихолинэстеразным препаратом
Абрамова Надежда Сергеевна
магистрант, Тюменский государственный университет, РФ, г. Тюмень
Дубровский Виталий Николаевич
научный руководитель, канд. биол. наук, доц., Тюменский государственный университет, РФ, г. Тюмень
В научной периодической печати имеются многочисленные данные указывающие на изменение активности транспортных АТФаз в условиях стресса. Также в литературе широко обсуждается вопрос о возможном влиянии холинреактивных систем на активность Na,K-АТФазы. В связи с этим целью данного исследования было изучение активности и магнийзависимых свойств Na,K-АТФазы в различных отделах головного мозга взрослых крыс линии Вистар в условиях действия иммобилизационного стресса после предварительной обработки прозерином (ингибитор холинэстераз).
Полученные данные показали, что активность Nа,К-АТФазы снижена в коре больших полушарий и хвостатом теле стрессированных животных. Снижение обусловлено возрастанием концентрации норадреналина, который подавляет действия фермента. Изучение магнийзависимых свойств показали, что при повышении концентрации ионов магния в инкубационной среде активность фермента падает, что связано с конформационными перестройками Nа,К-АТФазы. Также мы выяснили, что действие прозерина вызывает замедленый ответ катехоламинергической системы.
Активность ферментов, играет важную роль в развитии биохимической адаптации, обеспечивающая постоянство внутренней среды организмов в условиях внешней среды [1, с. 40]. Анализ фермента в разных тканях позволил выделить свойства Na,К-АТФазы не связанные с функцией транспортного насоса. Также фермент рассматривается как система рецепции и передачи сигналов в клетку, контролируемая холинреактивной системой [2, с.200; 3, с.47].
Воздействие стресса и холинреактивных систем могут оказывать большое влияние на активность транспортных АТФаз [4, с. 186]. Механизмы действия стресса проявляются рядом факторов, которые называются маркерами стресс-реакции, примером может служить концентрация аскорбиновой кислоты [5, с.102].
В связи с вышеизложенным нами была определена следующая цель работы: исследовать активность транспортных АТФаз в различных отделах головного мозга крыс, подвергнутых иммобилизационному стрессу и обработанных антихолинэстеразным препаратом.
В связи с поставленной целью были определены следующие задачи исследования:
1. определить концентрацию АК и ДАК в гомогенате надпочечников крыс, подвергнутых иммобилизационному стрессу и обработанных антихолинэстеразным препаратом;
2. определить активность Na,K-АТФазы в грубой микросомально-митохондриальной фракции коры больших полушарий, хвостатого тела и мозжечка головного мозга крыс, подвергнутых иммобилизационному стрессу и обработанных антихолинэстеразным препаратом.
Для количественного определения фракций аскорбиновой кислоты использовали метод Roe Kuethler, в модификации В. В. Соколовского с соавт. (1967) [6, с. 21]. Активность Na,К-АТФазы определяли по разности активностей между общей и Мg2+-АТФазой в инкубационной среде 1мМ уабаина. Содержание неорганического фосфата определяли методом Чена [7, с. 639]. Определение концентрации белка проводили по методу Лоури (Lowry et al., 1951) [8, с. 211].
Классическим косвенным маркером развития стресс-реакции является измерение в надпочечниках животных концентрации аскорбиновой кислоты[5, с. 105]. От концентрации аскорбиновой кислоты напрямую зависит количество норадреналина и адреналина. Таким образом, при снижении концентрации АК можно судить о развитиии стресс-реакции [9, с. 406]. Нами показано, что содержание АК в надпочечниках достоверно снижается по сравнению с контролем (табл. 1).
Таблица 1.
Концентрация аскорбиновой кислоты (мг%) в гомогенате надпочечников крыс, подвергнутых 40 мин иммобилизационному стрессу и обработанных прозерином
|
|
Аскорбиновая кислота |
Дегидроаскорбиновая кислота |
|
к |
1.771 ± 0.122 |
0.291 ± 0.046 |
|
с |
1.017 ± 0.028 *** |
0.006 ± 0.006 *** |
|
сп |
1.059 ± 0.036 *** |
0.132 ± 0 .017* |
Примечание: * – различие по сравнению с контролем статистически достоверно на уровне Р<0,05, *** – различие по сравнению с контролем статистически достоверно на уровне Р < 0,001, к – контроль, с – стресс, сп – стресс с прозерином
Полученные данные показали снижение фракций АК и ДАК в опыте с животными, подвергнутыми действию стресса и животных обработанных прозерином так как катехоламины являются основными гормонами стресса, то повышенный их синтез будет являться признаком развития у этих животных стресс-реакции. Менее выраженное снижение фракций АК в опыте с применением прозерина мы связываем с замедлением ответа со стороны катехоламинергической системы [10, с. 74; 11, с.1325; 12, с.269; 13; 376].
Изменение концентраций ацетилхолина и норадреналина в нервных клетках, может влиять на активность Nа+,К+-АТФазы. Так, понижение концентрации ацетилхолина и повышение концентрации норадреналина могут привести к снижению активности фермента [14, с.117]. Поэтому следующим этапом в работе стало измерение активности Nа,К-АТФазы в грубой микросомально-митохондриальной фракции коры больших полушарий, хвостатом теле и мозжечке крыс, предварительно обработанных и необработанных антихолинэстеразным препаратом - прозерином. Результаты исследования представлены (табл. 2).
Таблица 2.
Активность Na,K-АТФазы в грубой микросомально-митохондриальной фракции коры больших полушарий головного мозга, хвостатого тела и мозжечка крыс, подвергнутых стрессу и обработанных прозерином, при концентрации магния равной 3мМ в среде инкубации
|
Ткань |
Na,K-АТФаза |
||
|
к |
с |
сп |
|
|
КБП |
21,60±2,02 |
17,40±1,95* |
20,01±2,10 |
|
ХТ |
21,20±1,76 |
15,30±0,74* |
20,59±3,81 |
|
М |
19,71±2,03 |
19,63±2,98 |
22,56±3,34 |
Примечание: * – различие по сравнению с контролем статистически достоверно на уровне Р<0,05, к – контроль, с – стресс, сп – стресс с прозерином
Результаты показали, что активность Nа,К-АТФазы достоверно понижена в КБП и ХТ опытных животных, подвергнутых действию стресса по сравнению с контрольными группами животных, в то время как в фракции мозжечка, достоверных отличий активности фермента по сравнению с контролем не наблюдалось. Снижение активности Na,K-AТФазы в условиях стресса может быть связано с возрастанием концентрации норадреналина, который подавляюще действует на данный фермент. Отсутствие изменений в опыте с применением препарата может быть связано с накоплением ацетилхолина и менее выраженным возрастанием содержания катехоламинов, что имеет подтверждение в результатах исследования фракций аскорбиновой кислоты в гомогенате надпочечников [15, с. 161].
На следующем этапе работы нами была изучена активность Na,K-AТФазы в гомогенате коры больших полушарий головного мозга, хвостатом теле и мозжечке крыс в зависимости от содержания MgCl2 в среде для определения ферментативной активности (рис. 1–3).
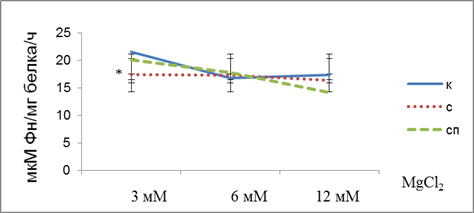
Рисунок 1. Активность Na,K-AТФазы в гомогенате коры больших полушарий головного мозга крыс, подвергнутых действию стресса и обработанных антихолинэстеразным препаратом в зависимости от содержания MgCl2 в среде инкубации. Примечание: к – контроль, с – стресс, сп – стресс с прозерином. * – различия с контрольной группой статистически достоверны при р <0.05
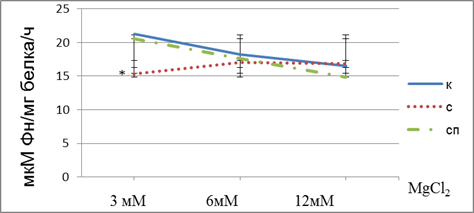
Рисунок 2. Активность Na,K-AТФзы хвостатого тела контрольной группы, крыс, подвергнутых действию иммобилизационного стресса и животных стрессированных с прозерином, в зависимости от содержания MgCl2 в среде инкубации. Примечание: к – контроль, с – стресс, сп – стресс с прозерином. * – различия с контрольной группой статистически достоверны при р <0.05
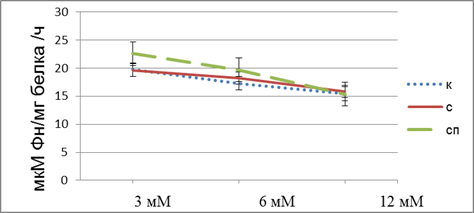
Рисунок 3. Активность Na,K-AТФазы в мозжечке головного мозга крыс, подвергнутых действию стресса и обработанных антихолинэстеразным препаратом относительно контрольной группы животных, в зависимости от содержания MgCl2 в среде инкубации. Примечание: к – контроль, с – стресс, сп – стресс с прозерином
Из представленных данных видно, что активность фермента снижается с ростом концентрации ионов магния в группе контрольных животных и животных, обработанных антихолинэстеразным препаратом. В то время как в группе животных, подвернутых действию иммобилизационного стресса рост концентрации ионов магния в инкубационной среде не приводит с соответствующему снижению активности фермента. Одной из причин нарушения зависимости активности фермента от концентрации ионов магния в среде для определения ферментативной активности может быть изменение его липидного окружения, вследствие активации перикисного окисления липидов, в связи с этим изменяется конформация фермента в мембране [16, с. 487; 17, с.1764]. Также причиной указанных изменений может быть увеличение в плазме крови животных, подвергнутых действию стресса концентрации эндогенные дигиталис-подобных факторов, которые обеспечивают поддержание гомеостаза в процессе адаптации организма к экстремальным раздражителям [15, с 162.].
Проведенные исследования позволили нам сделать следующие выводы:
Обнаружено снижение концентраций аскорбиновой и дегидроаскорбиновой кислот по сравнению с показателями контрольной группы в гомогенате надпочечников животных, подвергнутых действию иммобилизационного стресса, а также группе крыс, предварительно обработанных прозерином;
Показано снижение активности Na,K-АТФазы в коре больших полушарий головного мозга и хвостатом теле крыс в группе животных подвернутых действию стресса по сравнению с контрольной группой;
Отмечена измененная зависимость активности Na,K-АТФазы от концентрации ионов магния в среде для определения ферментативной активности в группе животных, подвернутых действию стресса.
Список литературы:
1. Архипенко Ю.В. Стрессорные повреждения ионных насосов миокарда и их адаптационная защита: дис., докт. биол. наук: 14.00.16 – патологическая физиология, 03.00.02. – М.: биофизика, 1992. – 42 с.
2. Болдырев А.А. Роль Na,K-насоса в возбудимых тканях. – М.: Международный Биотехнологический Центр МГУ им. Ломоносова, 2008. №3. – 206 с.
3. Васильева В.А. Содержание холестерина и аскорбиновой кислоты в надпочечниках в норме и при криптоспоридиозе. – М.: Фундаментальные исследования, 2007. №7. – 48–49 с.
4. Денисенко П.П. Роль холинергических систем в регуляторных процессах. – М.: Медицина, 1980. – 186–189 с.
5. Дубровский В.Н. Кыров Д.Н. Активность ацетилхолинэстеразы и содержание аскорбиновой кислоты в надпочечниках крыс при адаптации к иммобилизационному стрессу. – Т.: Фундаментальные исследования, 2004. №5. – 108 с.
6. Дубровский В.Н. Определение основных кинетических параметров холинэстераз из различных тканей крысы: учеб.пособие для практических занятий по биохимии. – Т.: Издатльствово Тюменского университетата, 2005. – 22 с.
7. Елаев Н.Р. Семенов Е.В. Изменение активности мембранных АТФаз мозга при воздействии холино - и адреномиметических веществ. – М.: Биохимия, 1974. – Т. 39, №. 3. – 636-640 с.
8. Заводская И.С. Морева Е.В. Фармакологический анализ механизма стресса и его последствий. – Л.: Медицина, 1981. – 213 с.
9. Лопатина Е.В. Пеннияйнен И.В. Крылов Б.В. Фармакологическая модуляция трансдукторной функции Na,K, ATФ-азы. – М.: Бюллетень экспериментальной биологии и медицины, 2008. – Т.146, №10. – 416 с.
10. Маслова М.Н. Активность мембранных ферментов эритроцитов при различных стрессорных воздействиях. – М.: Физиологический журнал имени И.М сеченова, 1994. – Т. 80, № 7. – 76 с.
11. Маслова М.Н. Молекулярные механизмы стресса. – М.: Физиологический журнал им И.М. Сеченова, 2005. – Т.91, №11. – 1320-1328 с.
12. Мосягин В.В. Влияние возраста и физиологического состояния животных на активность ферментных систем клеток, тканей и органов: дис., доктор биол. наук: 03.03.01, 03.01.04. – М.: Биохимия, 2011. – 273 с.
13. Розен В.Б. Основы эндокринологии. – М.: МГУ, 1994. – 384 с.
14. Силиванова Е.А. Влияние иммобилизованного стресса и внутримышечного введения неостигмина на вктивность ацетилхолинэстеразы и Na,K-АТФазы эритроцитов и головного мозга крыс: дис., канд. биол. наук: 03.00.04. – Т.: Издательство Тюменского государственного университета, 2006. – 126 с.
15. Соколовский В.В. Лебедева Л.В. Лиелуп Т.Е. Количественное определение фракций аскорбиновой кислоты. М.: Лабораторное дело, 1974. № 3. – 160–162 с.
16. Фролов Ю.П. Серых М.М. Макурина О.Н. Кленова Н.А. Подковкин В.Г Биохимия и молекулярная биология. – С.: Биохимия, 2004. – 501 с.
17. Chen P.S. Toribara T.Y. Warner H. Microdetermination of phosphorus Analyt Chem, 1957. – Vol. 28, – 1756–1758 p.
